Introduction
Creativity – the generation of products, works, or solutions that are both novel and useful (Hennessey & Amabile, 2010) – is perhaps humanity’s most distinctive trait, visible in domains as diverse as science and art, and distinguishing our earliest artifacts as seen, e.g., in the tool making and cave paintings of our ancestors. The relationship between intelligence and creativity has endured as a research topic (Guilford, 1967), partly because exceptional ability and creativity characterise the most impactful of human achievements (Cox, 1926; Murray, 2003), but partly too because the relationship has resisted resolution (Karwowski et al., 2016). As AI increasingly automates tasks requiring high cognitive ability, creativity-specific genetic variance may become increasingly valuable, driving innovation in fields like art, science, and entrepreneurship. Here, therefore, we investigate the genetic basis of creative achievement, as measured by the Creative Achievement Questionnaire (CAQ: Carson et al., 2005), and its association with the general factor of cognitive ability (g: Spearman, 1927) using a twin study. We first briefly review the literature on creativity and its proposed links to intelligence.
Creativity involves generating novel and valuable ideas or products, while intelligence entails learning and applying knowledge in new contexts (Gottfredson, 2002; Hennessey & Amabile, 2010). A longstanding debate questions whether creative achievement reflects general cognitive ability or a distinct mental process. Where cognitive ability tests involve items with single correct answers that can be deduced from available material, multiple creative outputs are possible from a given input and reproduction of existing solutions is not, by definition, deemed creative. Thus, despite the superficial similarities suggested by the involvement of novelty and utility in their definitions, the two constructs potentially differ in deep ways. Creativity, then, may be genetically distinct from intelligence, or no such dichotomy may exist, in which case creative responses would be simply a sub-case of the outputs from general learning and information-processing capacities core to cognitive ability (g).
Both views have been advanced. Early studies suggested intelligence and creativity were independent (Barron, 1963; Getzels & Jackson, 1962; Wallach & Kogan, 1965), but their limited scope and power left the relationship unresolved. Recent evidence indicates around 25% shared variance (Nusbaum & Silvia, 2011; Silvia, 2015). This new view suggests that learning and problem solving abilities such as spatial, verbal, and working memory used to deduce correct answers to problems (Raven et al., 1977) also promote creative achievement but fall substantially short of a full account of creativity. But the genetic underpinnings of creativity and its overlap with cognitive ability remain unclear.
Creative Achievement
The classic laboratory approach to measuring creativity was divergent thinking, such as generating unconventional uses for common objects (Guilford, 1956; Reiter-Palmon et al., 2019; Torrance, 1969; Wallach & Kogan, 1965). Jung and Hunter (2022) provide an excellent review of research and testing, including alternate uses tests, the requirement that these measures predict real-world creative achievement, and their discriminant validity from intelligence, ending with a call for more imaginative research into creativity. Divergent thinking was hoped to tap potential for creativity (Tinio & Barbot, 2017) but its validity has been subject to critique, with Warne et al. (2022) concluding that verbal and figural measures of divergent thinking do not form a coherent construct, while other research found that divergent thinking tests are heavily influenced by general cognitive ability, limiting their specificity as measures of creativity (Weiss et al., 2021).
Increasingly, researchers have shifted to using direct measures of creative achievement, prioritizing the face-validity and criterion validity inherent in real-world creative achievement as well as the bandwidth to include eminent creativity (Carson et al., 2005; Plucker, 2010). Here, we measure creative achievement with the Creative Achievement Questionnaire (CAQ: Carson et al., 2005). This reliable and valid instrument (Silvia et al., 2012) measures observable creative accomplishments across multiple domains (e.g., visual arts, music, scientific discovery; see Table 1 for full list). The CAQ can be scored as a total sum of achievements across domains or as the highest achievement level in any single domain, reflecting peak creative output. It is also often subdivided into artistic and scientific sub-scales. CAQ total scores correlated well with the personality trait of openness to experience (Kaufman et al., 2016; Silvia et al., 2009).
Comparing the relationship of cognitive ability and divergent thinking measures with creative achievement, a meta-analysis by Kim (2011) found modest correlations between creative achievement and both divergent thinking (r = .22) and IQ (r = .17), indicating that neither fully explains creative achievement. However, these phenotypic correlations are modest and do not address genetic architecture. This suggests that, at the least, cognitive ability leaves the bulk of creative achievement unaccounted for.
Genetics of creativity
Behavioural genetics provides a powerful tool to address questions regarding causal relationships between phenotypes (Martin et al., 1997; Martin & Eaves, 1977), allowing researchers to contrast models in which creative achievement is modelled as largely resulting from environmental contexts versus being heritable and models in which creative achievement is subsumed by cognitive ability, overlaps genetically or environmentally, or is quite independent of ability. Triangulation across distinct methods of genetic study supports a strong genetic basis for creativity. Roeling et al. (2017), for example, used profession (working in a creative profession versus not) as a phenotype for creativity in a twin study, finding that this was both strongly heritable (h2 = 0.70) and showed no evidence of effects of family environment. They noted also that such work is rare, with only 2-3% of people currently working in creative professions.
Molecular genetic studies, though in their infancy for creativity, are now yielding polygenic scores – scores which aggregate the effects of multiple genetic variants – explaining 1-2% of variance in phenotypes such as rated occupational creativity (Li et al., 2024). Other molecular work has used aesthetic chills – a specific marker of openness (McCrae, 2007) – showing that this too exhibits a statistically significant molecular genetic signal (Bignardi et al., 2025). This work is unfortunately not yet far enough advanced to clearly rule in or out a strong genetic overlap of creativity with intelligence.
de Manzano and Ullén (2018) advanced the field with a twin study examining intelligence, openness to experience, and creative achievement (CAQ, adapted to seven domains: five artistic, two scientific, see Table 1). They found modest phenotypic overlap among these traits, with substantial unique genetic influences on creative achievement, particularly in artistic domains. Their findings suggest that creativity is not fully explained by cognitive ability or openness, supporting a hybrid model of genetic influences with modest phenotypic overlap between intelligence, openness, and creative achievement. Most (~90%) of the shared phenotypic variance was due to genetic effects. Alongside this however, both artistic and scientific maximum creativity showed substantial unique genetic influence which could not be reduced to either openness or cognitive ability. In terms of the amounting of phenotypic variance of intelligence and creative achievement, around 33% was shared for artistic achievement, rising to 58% for scientific creative achievement. Cognitive ability had stronger effects on scientific creative achievement compared to artistic achievement (standardized path coefficients of .48 versus .28), while openness to experience showed stronger genetic effects on artistic creative achievement than on scientific creative achievement (.62 versus .54 respectively).
Models and Hypotheses.
Building on existing work, we undertook a twin study in which we assessed cognitive ability and creative achievement (CAQ) in twins, using the full CAQ and extending it from the existing two domains of art and science to a domain of action, previously not examined, but which is notable for creative achievement, namely in entrepreneurial, military, professional and political achievement. The twin data and three phenotypic measures of each of creativity and cognitive ability allow a test of contrasting models of genetic and environmental overlap of the cognitive ability and traits in latent models.
Based on findings of significant evidence for heritability of creativity, and phenotypic evidence for partial overlap of cognitive and creative processes, and twin study support for an overlap model, differing effects for scientific/inventive and artistic creative achievement, we postulated three competing models (see Table 2). Finally, in addition to these three hypotheses, we explored if the best fitting model differed for CAQ sub-scales of scientific versus artistic creativity (de Manzano & Ullén, 2018).
Method
Participants
The sample was drawn from the Brisbane Adolescent Twin Study (Couvy-Duchesne et al., 2018; Mitchell et al., 2019). This sample was initially recruited through Queensland schools, via the Australian Twin registry and through word of mouth Gillespie et al. (2013). All participants were volunteers. A total of 2,141 twins from 549 MZ pairs and 822 DZ pairs (391 opposite-sex and 431 same-sex) completed the ability test: Male = 803; Female = 1,338, Mean age 28.62 years (SD= 7.32). Complete data for both the ability and the CAQ tests (administered in separate test blocks) were available for 1,622 individuals. For the twin modelling descriptive data, see Table 3 showing mean, standard deviation, and twin-pair correlations by zygosity). Zygosity was determined by DNA genotyping of highly polymorphic DNA microsatellite polymorphisms and the X/Y amelogenin marker for sex determination (Wright & Martin, 2007).
Measures
Creative achievement was measured using the full CAQ (Carson et al., 2005). The CAQ is reliable, with a mean coefficient alpha (internal consistency) reliability of 0.77 (Yörük & Sen, 2022). Following precedent in creativity research (e.g., Plucker, 2010), we added a third “active” domain to include creative accomplishments in business, the military, and politics. These capture real-world creative achievements which fall outside the arts yet express creativity including developing products and services, strategy, and policy. For analysis, we follow the approach of de Manzano and Ullén (2018) and scored the CAQ as the maximum accomplishment achieved across each domain, and in total across all domains. Three subscales were scored for analysis: Artistic (summing Visual Arts, Dance, etc.); Scientific (Scientific Discovery, Inventions), and Action/Enterprise (Entrepreneurship, military, political) using the specialties listed in Table 1.
Cognitive ability was measured using three sub-tests. Eight items were selected from the open science International Cognitive Ability Resource (ICAR: Condon & Revelle, 2014). Four ICAR items assessed Numeric puzzles (ICAR numeric). A further four assessed Letter number sequence deduction (ICAR seq). Vocabulary was measured using 10-items based on the General Social Survey vocabulary measure, delivered in a multiple-choice format with a target word (e.g., EDIBLE) and five multiple choice options (e.g., auspicious; eligible; fit to eat; sagacious; able to speak) and subjects were to pick the word closest to the meaning of the target. These measures were chosen for their open-access availability, reliability given the brevity of the scales, and coverage of fluid and crystallized intelligence (Condon & Revelle, 2014). Omega-t reliabilities (ω-t) for the Number, Sequence, and Vocab scales were .55, .67, and .64, respectively.
Procedure
The research had ethical approval from University of Edinburgh and QIMR Berghofer Medical Research Institute ethics committees. Subjects were contacted to recruit them, and after consenting, completed the study online using the widely used and robust Qualtrics.com web-based survey database platform, embedded in a larger block of testing for other projects. Participants completed the assessment in one session, beginning with the cognitive ability tests, and an effort measure, followed by the CAQ. To ensure participant confidentiality, data are anonymized using unique IDs. No personally identifiable information was collected, and the researchers had no access to individual participant identities during or after the data collection process.
Statistical Analyses
All statistical analyses were completed in R (R Development Core Team, 2025), OpenMx (Boker et al., 2011; Neale et al., 2016) and umx (Bates et al., 2019; Castro-de-Araujo et al., 2026).
Data were analysed using models based on the classic twin design (Jinks & Fulker, 1970; Neale & Cardon, 1992). These models use the fact that identical (monozygotic: MZ) twin pairs share 100% of their genetic makeup while fraternal or dizygotic (DZ) twin pairs share on average 50% of their genetic makeup. Data from MZ and DZ twin phenotypes can thus be decomposed into independent additive genetic (A), shared environmental (C), and non-shared or unique environmental (E) components. More complex family designs allow estimating dominance effects and shared environment simultaneously. Modern twin modelling utilises the extended structural equation model (SEM) framework, permitting complex multivariate designs to be tested (Eaves et al., 1978; Neale & Maes, 1996). These models were estimated from raw data using full-information maximum-likelihood (FIML) analysis. Nested models were compared using chi-square (χ²) tests of the change in minus log-likelihood (Δ-2LL), with degrees of freedom equal to the difference in estimated parameters. The -2LL χ² test is employed to compare the fit of nested models, particularly in genetic epidemiology. This test assesses whether a more constrained model (e.g., one dropping genetic influences) fits the data significantly worse than a less constrained model (e.g., one allowing for genetic effects). The change in log-likelihood test statistic (Δ -2LL) is asymptotically distributed as a chi-square when the null hypothesis is true, with degrees of freedom equal to the difference in the number of parameters estimated between the two models.
We first evaluated univariate heritability of each scale using ACE models, followed by multivariate models of the ability and CAQ scales, testing competing predicted common pathway structures (see below). A common pathway model posits that a latent factor mediates genetic and environmental influences on multiple traits, reflecting shared underlying mechanisms, this allows modelling of variance in multiple traits simultaneously, while respecting the theoretical dependence of these traits on more general latent factors (Kendler et al., 1987; Martin & Eaves, 1977). The common pathway model uses one or more common latent factors each acting as a focal point of causation, mediating additive genetic (A), shared environmental (C), and non-shared environmental (E) influences on the measured variables. As such, these latent common factors correspond to a mechanism through which genetic and environmental effects act. The common pathway, thus, will fit poorly if such a coherent psychological architecture does not exist. For multivariate modelling, ACE models were used as the saturated baseline model against which common pathway models specified to implement the three theoretical models were evaluated. A correlated-factors model was also fit as a baseline model, and gave comparable results, suggesting that the ACE model could be used as a valid baseline (Verhulst et al., 2019).
Because CAQ scores typically exhibit positive skew, we conducted a sensitivity analysis using data-driven Yeo-Johnson power transformations (Yeo & Johnson, 2000). Refitting our structural models using these mathematically optimized variables yielded substantively identical heritability profiles to our raw analysis (e.g., heritability (a) of creative achievement shifted negligibly from .56 to .58), confirming that our FIML estimations are robust to the observed skewness and capture genuine latent genetic architecture.
Results
Descriptive statistics for the creativity measures are shown in Table 3. Intraclass correlations of MZ and DZ twins for each measure suggested evidence of heritability for all scales. We also tested initial univariate models of heritability for each trait in a structural modelling framework using univariate ACE models, which decompose the variance of the observed traits in twin 1 and twin 2 into additive genetic (A), shared environmental (C), and unique environmental (E) components (Neale & Cardon, 2013). Prior to analysis, age was regressed from the twin data (McGue & Bouchard, 1984). The results of these models are shown in Table 5 in the columns labelled a, c, and e.
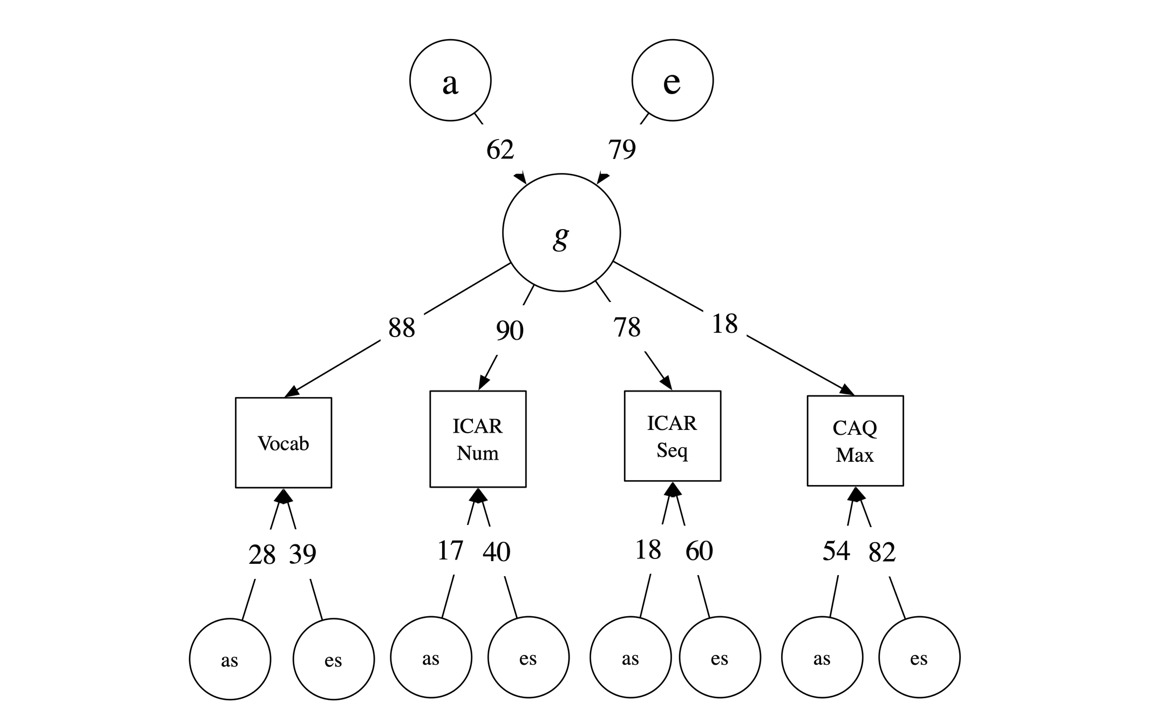
The first theoretical model evaluated was a single-factor common-pathway model of the three cognitive measures and the maximum score on the CAQ (see Fig. 1). This model which imposes a single common factor which loads on all measures, allows us to assess the extent to which all four measures can be treated as indicators of a common general or g-factor of cognitive ability. This common pathway model itself was first compared to a saturated model (four-factor ACE model) to assure that the one common factor model fit adequately, before evaluating our more specific hypotheses about this predicted model by path removal and significance testing. As can be seen (Table 6) the one-common-factor model fit adequately. We also assessed the effect of shared environment in this model as these were all estimated at or near zero. All C and cs effects could be dropped without loss (p = .970, see Table 6). Having established that a 1-common-pathway model with no shared environment effects fit well, we evaluated simplifications of this model to assess our empirical hypotheses.
We first tested if CAQ scores could be modelled as having no genetic or environmental links to this g-factor by testing deletion of the path from the latent g-factor to CAQ. As shown in Table 6, this model proposing no impact of general ability on CAQ fit very poorly (χ²(1) = 12.71, p < .001). This supports the need for influences on CAQ from the general factor. Next, we tested if the residual specific heritable variance (as) on CAQ could be dropped. A large residual genetic effect would support CAQ representing a separate causal process distinct from general ability. This CAQ-specific genetic influence could not be dropped without highly significant loss of fit (χ²(1) = 84.85, p < .001). Moreover, the magnitude of the specific genetic path to CAQ (a = .70; see Figure 1) was larger than its loading on the g-factor (.18), implying that most of the genetic influence on creative achievement is independent of general ability. Figure 1 illustrates this final one-factor common pathway model, showing a general cognitive factor (g) loading on all measures, with CAQ exhibiting substantial unique genetic variance independent of g.

This one factor model of creative achievement showed that creative achievement is heritable, that it has modest (but significant) influences from general cognitive ability, that there was evidence for a large element of genetic influence on creative achievement distinct from cognitive ability and specific to creative achievement, and, finally, that shared environment had non-significant and negligible effects on either cognition or creative achievement.
We next modelled the artistic, scientific, applied outputs of creativity as distinct forms of creativity. To this end, a second model was built incorporating the full science and arts sub scores of the CAQ and the new political, military and entrepreneurial achievement measures.
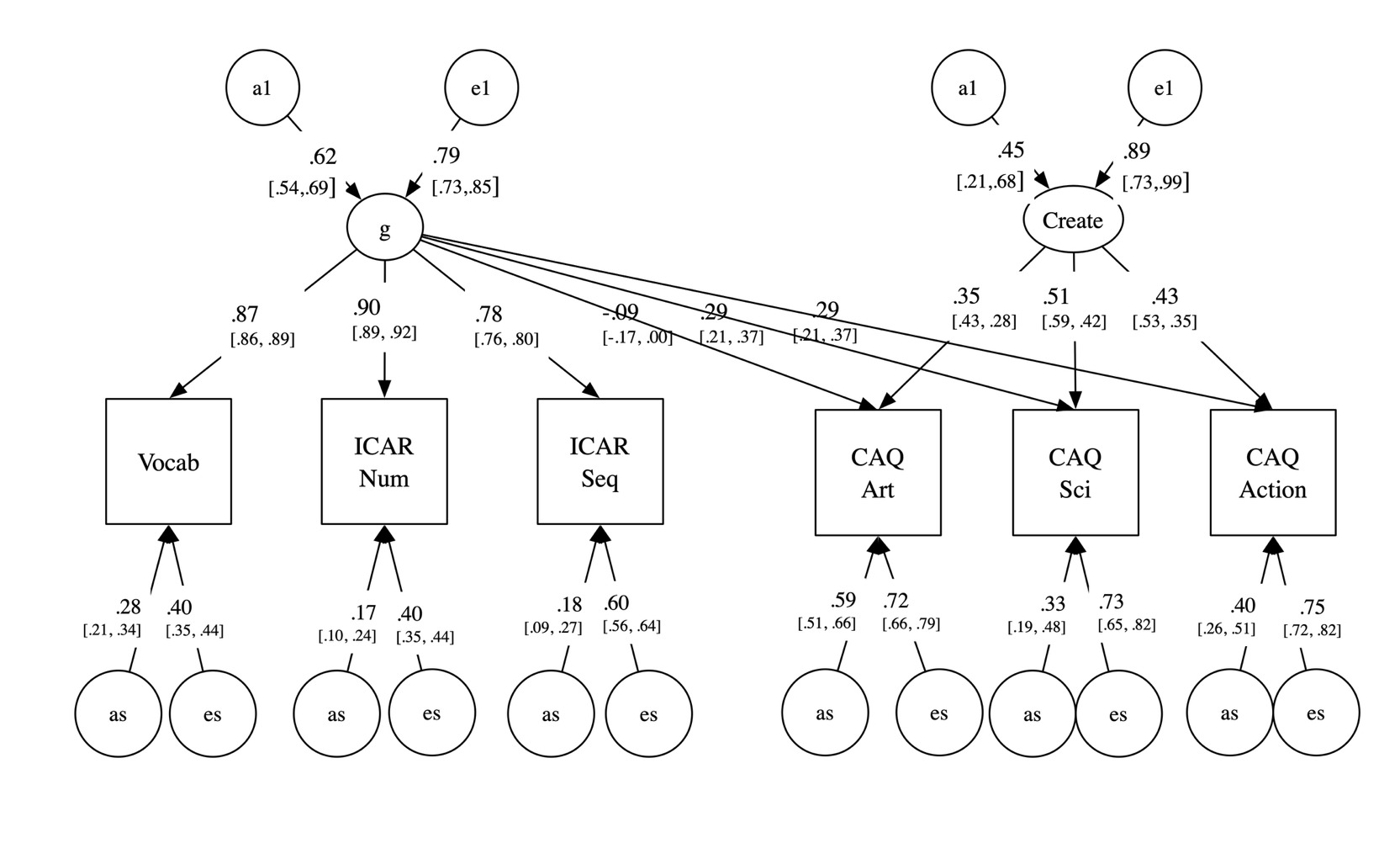
One and two common pathway models were tested. A one common pathway model did not fit well relative to the ACE baseline. However, a two common pathway model did fit well (see Table 7). The first common factor was general, loading on all tests. The second common-pathway latent variable loaded on the creativity measures, i.e., it was a latent trait of creativity. Shared environment effects were estimated at or near zero and could be dropped from the model without loss of fit (χ²(8) = .00, p = 1.00).
Taking this model with shared environment dropped as a new baseline, we next tested if the creativity factor was distinct from cognitive ability. This was done by setting the loadings of the creativity latent factor onto the cognitive ability measures to zero. This could be done with no significant loss of fit (χ²(3) = 2.067, p = .559 model “Independent Creativity Factor”). The non-significance of the test for this model confirms the hypothesis that factor two is specific to creative achievement alone and has no effect (positive or negative) on ability. We then attempted to drop loadings from the g factor onto creative achievement. As can be seen in Table 7, this was not possible for science or enterprise, and was marginal for art. The AIC (Akaike Information Criterion: Akaike, 1974) index of the relative quality of statistical models was higher for a model dropping loadings from the g factor onto artistic creative achievement. As this criterion assesses how well a model fits the data while penalizing for complexity, with a lower AIC score indicating a better model that loses less information, it suggests that the best model among the candidates for this dataset would include an influence of cognitive ability on artistic creative achievement.
The fit statistics for this final model, with no shared environment, independent, heritable latent factors of general ability and creative ability is shown in Table 7 and presented graphically in Figure 2. Critically, the genetic correlation between the general cognitive factor and the latent creativity factor was exactly zero. Genetic influences on creative achievement therefore operate through a distinct pathway that shares no genetic variance with g. Small residual genetic paths from g to the observed CAQ variables remained (.29 for science and action; –.08 for art), indicating modest domain-specific genetic overlap at the indicator level only
_and_latent_creativity_(create.png)
Discussion
The most striking finding is the complete genetic independence of the latent creativity factor from g (rg = 0). This demonstrates that the genetic architecture of real-world creative achievement is not merely a downstream consequence of general intelligence but reflects a separate, heritable system that operates across artistic, scientific, and enterprising domains. Our best fitting twin genetic model revealed the need for two latent factors, with no genetic correlation between the latent creativity factor and g and with g impacting creative achievement via smaller observed-level causal paths (see Fig. 2). The results support the hypothesis that creative achievement is heritable, is independent of cognitive ability at a latent level, but that cognitive ability directly raises specific achievements, especially scientific and enterprising. This also highlighted the value of distinguishing between achievement in the sciences and in the arts. Moreover, the results validated our extension of the creative realm of the CAQ into business, politics, and military enterprise. These valuable domains should be included in future measures. Associations with cognitive ability were higher for STEM achievement than art. One possibility for this is that technology offers more opportunities for production which does not require generating a creative hypothesis oneself. A distinct hypothesis is that technology’s greater dependence on mathematical models raises the bar in terms of IQ for productive involvement in science. This can be evaluated.
Our findings align with and extend those of de Manzano and Ullén (2018), who also used a twin study to explore the genetic basis of creative achievement (CAQ) alongside intelligence and openness to experience. Like their study, we found high heritability for creative achievement and substantial unique genetic variance, not explained by general cognitive ability, supporting a hybrid model. However, our study advances their work by incorporating a full and broader CAQ (including business, military, and political domains) and using a three-factor common pathway model, which revealed a distinct creativity-specific latent factor loading on both artistic and scientific achievement. Like de Manzano and Ullén (2018), we found nominally higher effects of cognitive ability on scientific compared to artistic achievement (r = 0.48 vs. 0.28 for artistic in their study, and for us .29 and a (non-significant) -.08 path to art).
Collectively, the findings underscore the creativity-specific genetic mechanisms, and the need to understand these better, potentially through molecular genetic studies building on well powered datasets. Together with the results of de Manzano and Ullén (2018), the present results support the idea that cognitive ability is causally linked to increased creative achievement, but highlight the large fraction of genetic influences on creative achievement that are specific to creativity, accounting for most of creative achievement. Future work using very large datasets, perhaps by capitalizing on imputed measures of creativity, but preferably including the CAQ in a large molecular study would be a useful start to understanding this creativity-specific system (Roeling et al., 2017). Work on Openness to Experience will also be invaluable, as this is the most promising human drive to generate novel ideas (Jung & Hunter, 2022).
The emergence of a distinct creative achievement factor encompassing entrepreneurial, military, and political domains suggests that creativity research has historically under-sampled specific vocational interest profiles. Mapping these domains onto interests (Rounds & Su, 2014) reveals a clear alignment: while traditional CAQ scales capture ‘Artistic’ (expressive, unstructured) and ‘Investigative’ (analytical, scientific) drives, our extended domains capture ‘Enterprising’ creativity—the drive to strategically organize people and resources to achieve novel organizational or societal outcomes. This Enterprising facet of creativity is highly heritable and distinct from general cognitive ability, highlighting the need to broaden our conceptualization of where human creativity is deployed in the real world across Artistic, Investigative and Enterprising interests.
Limitations and Future directions
It would be valuable to replicate and extend the research to independent samples. Openness to experience was not measured in this study. Our results pave the way for future research into the molecular genetics of creativity and its implications for education, innovation, and AI-driven societies. The results support the use of the CAQ in genetic research, and incorporating it, for instance, into large biobanks is an obvious extension and enhancement of work initiated by Li et al. (2024) in generating genetic polygenic scores and causal genetic variants underlying elevated creativity. Integrating personality traits like openness to experience and leveraging large-scale biobanks will further elucidate the mechanisms underlying human creative potential. Openness is related to creativity (Oleynick et al., 2017), contributes genetically to creative achievement (de Manzano & Ullén, 2018), and has plausible links to artistic and technological expressions of creativity via its aesthetic and intellect facets respectively (Kaufman et al., 2016) (though note that the “intellect” elements of openness appear to reflect confounding with cognitive ability (Gignac et al., 2020). Adding a measure of openness would allow further genetic insight into this important but neglected trait. There is a need for formal psychological models explaining the modest links of creative achievement with cognitive ability (Nijstad et al., 2010). As achievement involves goal-setting (Bates et al., 2023) and persistence (Bates, 2024), incorporating conscientiousness in models of creativity alongside openness would also be valuable (Jauk et al., 2014). The result suggests that creativity is expressed in scientific/technological, artistic and, also, in domains of entrepreneurial enterprise. It is important to understand why cognitive ability does aid creativity but does not explain all of it. Why not? There are exciting pointers in this direction from computer science as AI researchers grapple with creativity (Kamb & Ganguli, 2024).
Conclusion
This twin study demonstrates that creative achievement is highly heritable but genetically distinct from intelligence at the level of latent constructs. g accounts for approximately 10% of genetic variance in creative achievement; the remainder is creativity specific. It extended the CAQ to include entrepreneurial/active achievement as well as science and arts. These findings highlight that creativity is not restricted to art. This hybrid model of cognitive ability and creativity-specific genetic and environmental factors highlights the need to explore the role of both in human accomplishment.