Introduction
From the extensively analyzed cognitive and non-cognitive traits, general cognitive ability (GCA or g), as measured by standardized tests, shows the most outstanding stability values across the life span (Haier et al., 2023). Evidence obtained from the 1932 and 1947 Scottish Mental Surveys (SMSs) provides a sound example. Findings reveal stability values from early adolescence (age 11) to old age (> 70 yrs.) ranging from 0.73 to 0.78 (Deary, 2014). The implication is that individuals keep their relative ranks within the cognitive ability distribution as they age. Cognitive ability scores do change across the life span, but these changes are far from random. Further research results from the SMSs support the conclusion that DNA differences drive cognitive stability across the life span to a meaningful degree, whereas non-genetic (environmental) factors drive the identified cognitive changes (Deary et al., 2012).
Meta-analytic results after the consideration of fifteen longitudinal twin and adoption studies are consistent with the SMSs’ findings just described. These are the main conclusions after analyzing data for individuals ranging from age 0.5 to age 77. First, cognitive ability differences show low stability in early life (0.30). Second, these stability values show a steady increase as individuals age (0.6 at age 10 and 0.7 at age 16). Third, this growing trend reaches asymptotic values of 0.78 in early adulthood. Fourth, genetic factors do not account for cognitive stability in early life, but they show increased relevance as individuals age reaching a contribution of 0.65. Fifth, shared environment shows a contribution of 0.24 to cognitive stability in early life but fades away with aging. Finally, non-shared environment increases its contribution to cognitive stability across the life span reaching a value of 0.20 in adulthood. These findings support the statement that genetic factors are behind increased cognitive stability as individuals age (Tucker-Drob & Briley, 2014).
The observed changes in cognitive stability across the life span are highly relevant for the trust we usually confer to the premature identification of high cognitive ability children (aka. intellectually gifted). If cognitive stability is low at early ages, then substantial changes can be expected in the children relative ranks, within the cognitive ability distribution, as they age. Importantly, unreliable early identification of high cognitive ability children might have undesired consequences. As underscored by Asbury and Plomin (2014), early labels attached to terms such as ‘intellectual giftedness’ are usually detached from performance at later ages. Gifted children at, say, age 4 or 6 are erroneously thought to keep their high levels irrespective of the changing maturation process. On the other side of the coin, early labels attached to terms such as ‘non-gifted’ preclude later identification because they are erroneously thought to remain at their cognitive levels. As a result of both presumptions, gifted and normative classrooms comprise children improperly identified. It is important to acknowledge that remarkable changes should be expected because, as noted above, genetic factors are much less relevant than non-genetic factors, such as home learning, when accounting for cognitive ability differences measured at early ages. Therefore, the relative standing of high ability and normative children in the preschool years is expected to change as they age. Paying attention to individuals showing high cognitive ability is relevant both at the personal and social levels (Lubinski, 2016), but the tendency to premature identification without proper follow-ups might lead to troubling outcomes. Uncovering and quantifying the expected changes is needed for a cautious and responsible approach. Furthermore, obtained results might be of great help to establish the best time window for achieving a reliable identification.
Research supports the latter cautionary notes (Kawakami et al., 2024). After the analysis of the educational achievement (EA) trajectories from age 7 to age 16 of 4,175 participants in the Twins Early Development Study (TEDS), findings reveal that students gravitate to their genomically predicted levels as they age. Those with better academic marks than expected (according to their genetic potential) at early ages (overachievers) showed a decrease in their scores over time, whereas those with worse scores than expected at early ages (underachievers) showed an increase in their scores. At age 16 students regressed to 38 % of the genomic predicted EA levels, on average, and most of these changes were noteworthy at age 9. Importantly for the main research purpose of the present study, non-genetic factors such as students’ family SES were unrelated in the long run with age-related changes in EA.
Here we pursue this main research question: which personal and situational factors distinguish children showing high cognitive ability at early age (4 and 7 yrs.) and keeping their high scores at later ages (12, 16, and 21 yrs.) from those who showed high ability at early age but lose their top levels afterwards? This key question will be complemented by the analysis of those showing normative ability at early ages for identifying those moving upwards and downwards or keeping their levels. The answer to this secondary question will allow knowing if the operating factors identified for the main question also hold for the normative band of the cognitive distribution.
We hypothesize that if developmental changes in the participants’ cognitive ability levels run in tandem with their spontaneous brain maturation processes (linked to their genomes), then the situational factors (schools and families) measured across time (from age 4-7) will not predict the expected upwards and downwards cognitive changes across age (12, 16, and 21 yrs.). Testing this hypothesis requires several steps. First, identifying individuals with high and normative general cognitive ability scores at age 4 and age 7. Second, characterizing their developmental trajectories overtime. Third, evaluating the association between the change in cognitive ability and variables such as family socioeconomic status (SES), home environment, school engagement, behavior problems, and life events. Finally, evaluating the association between individuals’ polygenic scores with their changes in cognitive ability.
Method
Dataset
For this study, we analyzed data from participants in the Twins Early Development Study (TEDS). TEDS is one of the world’s largest twin cohorts, investigating how genetic and environmental factors shape individual differences in cognitive and learning abilities, behaviors, and emotions in the context of typical development (Lockhart et al., 2023; Rimfeld et al., 2019). The dataset for the current study comprised 11,119 individuals. From this dataset, we selected two groups of children considering their scaled scores in general cognitive ability (GCA) at age 7, GCA100 and GCA115. GCA100 comprised individuals scoring between 99 and 115 (99 < GCA < 115), whereas GCA115 comprised individuals scoring above 115 (GCA > 115). The latent curve models (LCM’s) considered in the present study applied a full information maximum likelihood (FIML) method, allowing to examine the available data for 3,958 individuals in the GCA100 group, and 1,580 individuals in the GCA115 group.
Measures
Table 1 summarizes the measures in the present study. Table S1 shows the descriptive statistics of interest. Three measures were obtained at different ages and hence they were time-variant: general cognitive ability (age 4, 7, 12, 16, and 21), home environment (age 4, 7, 12, and 16), and behavior problems (age 4, 7, 12, and 16). Six measures were time invariant because they were obtained at one single time-point: two polygenic scores, socioeconomic status (age 7), home chaos (age 12), school engagement (age 16), and life events (age 21). We provide brief descriptions of these measures next (see Supplementary Material for further details).
General cognitive ability (GCA) is the dependent variable of interest. Because different specific measures were administered at different ages, we computed a general ability composite. Research shows that estimates of this general ability capture the reliable predictive validity of the cognitive ability construct (Watkins & Canivez, 2021; Zaboski et al., 2018). Two polygenic scores were selected from the complete dataset. One was derived from the Lee et al.'s (2018) study, whereas the other was computed after the Okbay et al.'s (2022) study. Home environment (parental discipline) was assessed by the structured interview of Deater-Deckard et al. (1998) and by The Confusion, Hubbub and Order Scale (CHAOS). Behavior problems were quantified by the Strengths and Difficulties Questionnaire (Goodman, 1997). Family socioeconomic status (SES) was computed after the educational and occupational qualifications of both parents. School engagement was measured by the School Engagement scale (Appleton et al., 2006). Finally, life events were measured by the Coddington’s (1972) scale.
Home environment and behavior problems were time variant predictors of general cognitive ability (GCA), while polygenic scores, SES, home chaos, school environment, and life events were time invariant predictors. Importantly, only measures obtained before the criterion measure were considered predictors, except for the life events measure, which comprises memories from past events throughout the observed period. This is further specified in the next section.
Data Analyses
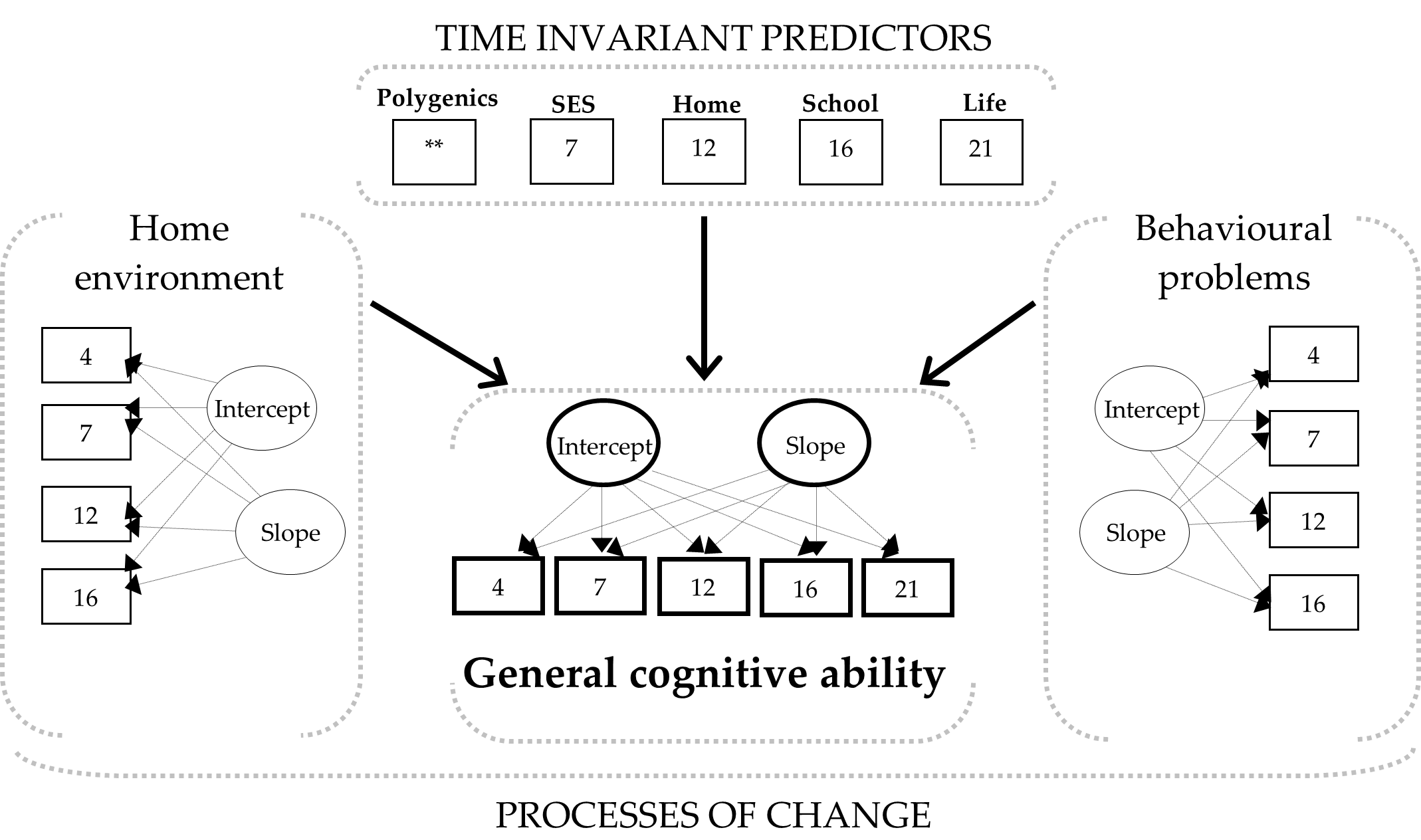
Figure 1 shows an overview of the analytic approach undertaken in the current study. There were three processes of change: general cognitive ability (GCA), home environment (HE), and behavioral problems (BP). Time invariant predictors included polygenic scores, socioeconomic status (SES), chaos at home (evaluated by parents and children), school environment, and life events. We specified latent curve models (LCMs) in two stages, unconditional and conditional (Curran & Hussong, 2002; Preacher et al., 2008; Willet & Sayer, 1996).
The unconditional latent curve models (LCMs) served to examine the three processes of change in GCA, HE, and BP. For each process, the measures obtained across age were thought to change based on two latent factors, intercept and slope. The intercept factor represents the respective measurement at the start (age 7) or at the end of the considered time window (age 16 or age 21). The slope factor represents the rate of change per time point during the observed period.
The conditional LCMs allowed to analyze the interrelationships among the processes of change and the time invariant predictors to account for the observed changes in GCA. Within the conditional LCMs, GCA was specified as the criterion process of change (intercept and slope), whereas the change in HE and BP processes (intercepts and slopes), along with the time invariant predictors, were specified as the causal factors of the changes observed in GCA. As noted above, the parameters of interest for the specified models were estimated with the full information maximum likelihood method (FIML) (Bollen, 1989).
.png)
The goodness of fit was evaluated with the chi-square value and p-value, the comparative fit index (CFI), the Tucker-Lewis index (TLI), the root mean square error of approximation (RMSEA), and the standard root mean residual (SRMR). Appropriate model fit holds with lower non-significant chi-square values, CFI and TLI closer to 1, and values below .08 for the RMSEA and SRMR indices (Hu & Bentler, 1999). Model fit was assessed comprehensively using all these indices together rather than using a single index. The statistical analyses of these models were conducted with the lavaan package from the R software, version 3.5.0 (R Development Core Team, 2014; Rosseel, 2012).
Transparency and Openness
This study was preregistered in OSF (https://osf.io/zjfd2/) The data used in this study were obtained from the Twins Early Development Study (TEDS). Researchers can apply for access to TEDS data by signing a Data Access agreement. The full data access policy and procedures are described in https://www.teds.ac.uk/researchers/teds-data-access-policy. The corresponding author will share the R code used upon request.
_at_the_individual_level_within_the_.png)
Results
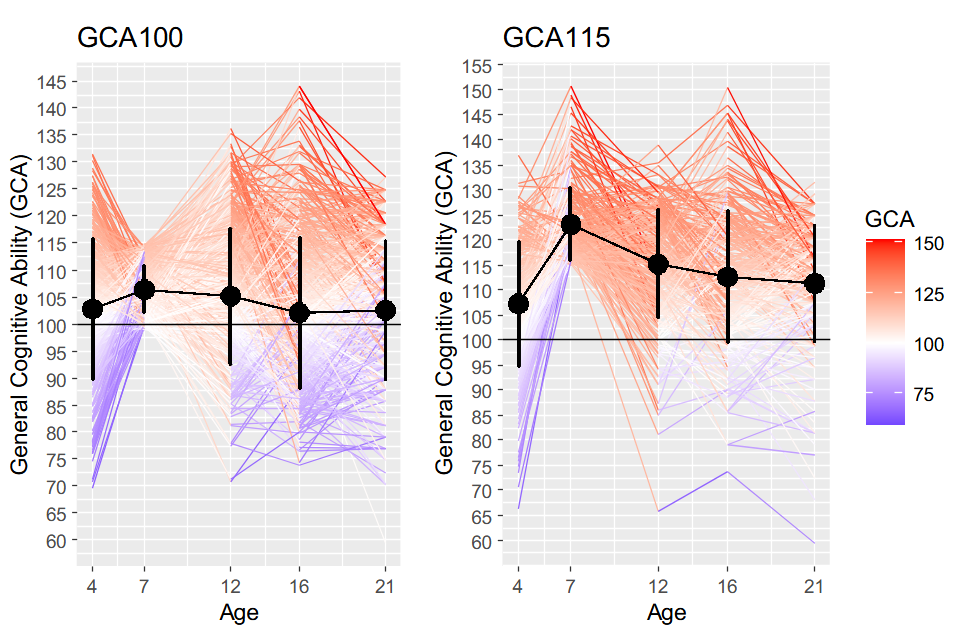
Figure 2 shows the changes in general cognitive ability (GCA) between age 4 and age 21 for both groups, GCA100 and GCA115. There was a remarkable variability regarding individual trajectories. Some individuals increased their scores, while others decreased from age 7 onwards. Individuals with the highest scores at each time point had the larger decrements at the next time point. Conversely, individuals below the mean (100) at ages 12 and 16 went up again at subsequent ages.
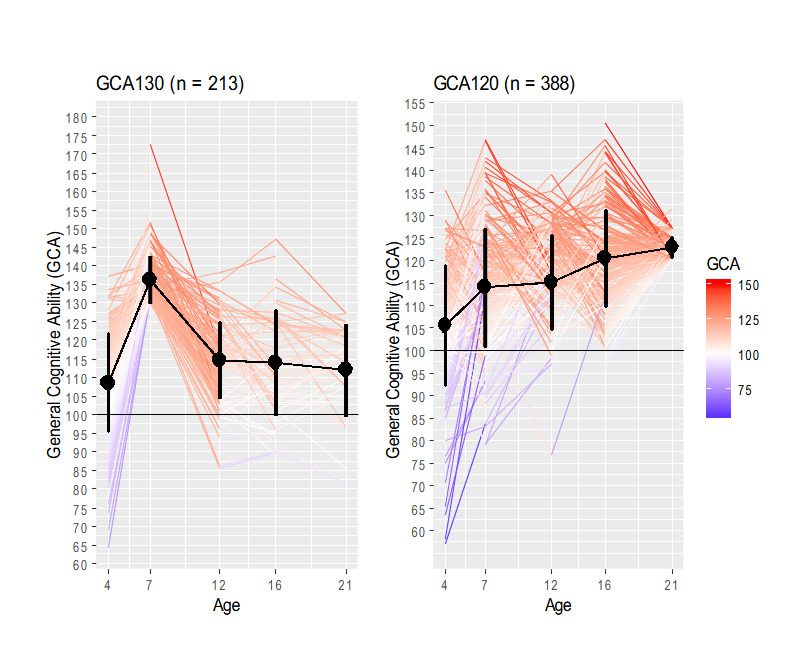
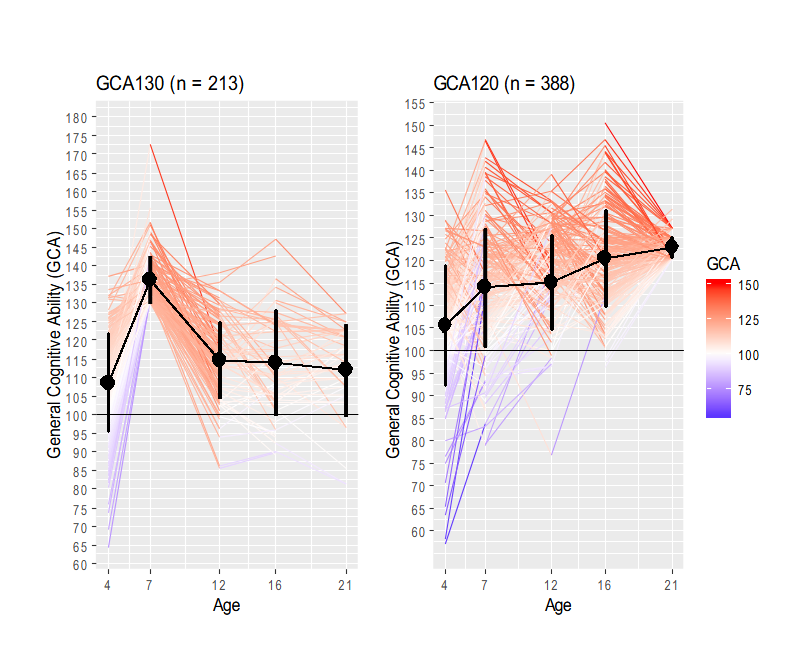
Furthermore, children scoring above 130 at age 7 (n = 207) and children scoring above 120 at age 21 (n = 388) show complementary trajectories (Figure S1). Most children scoring above 130 at age 7 lost their values afterwards whereas those scoring above 120 at age 21 came from a wide range of GCA scores. When the focus is placed at the early age of 7 years old, the average trend stabilizes from age 12 (left panel of Figure S1). On the other hand, when the focus is placed at age 21 there is a broad range of cognitive ability and an upward average trend starting at age 12 (right panel of Figure S1).
Unconditional GCA model
The unconditional LCMs outcomes for GCA shown in Table 2 reflect the noted intraindividual variability over time. The unconditional model outcomes for the process of change in home environment (HE) and behavioral problems (BP) are shown in the supplemental material (Table S2). The unconditional model fit outcomes are also shown in the supplemental material (Table S3). The outcomes from the model fit evaluation support that the unconditional models provided an appropriate representation of the observed data. Even though the chi-square were highly significant, the rest of the fit indices (CFI, TLI, RMSEA, and SRMR) yielded values within the expected range of goodness-of-fit.
Table 2 shows the parameter estimates for the unconditional LCMs in the process of change for general cognitive ability (GCA). For both groups, GCA100 and GCA115, their mean scores at age 7 were 106 and 118 points, respectively. Mean scores decreased progressively reaching values of 102 for GCA100 and of 108 points for GCA115 at age 21. These decrements are summarized by the mean slopes, which indicate a statistically significant mean decrease of -.29 (GCA100) and -.73 (GCA115) per time point. The variability associated to the mean scores at each point (Sd) increased progressively and quite similarly for both groups. Furthermore, there was statistically significant variability associated to these mean values regarding the slope (rate of change) of .91 and .86, highlighting the quite heterogeneous trajectories in GCA shown in Figure 2.
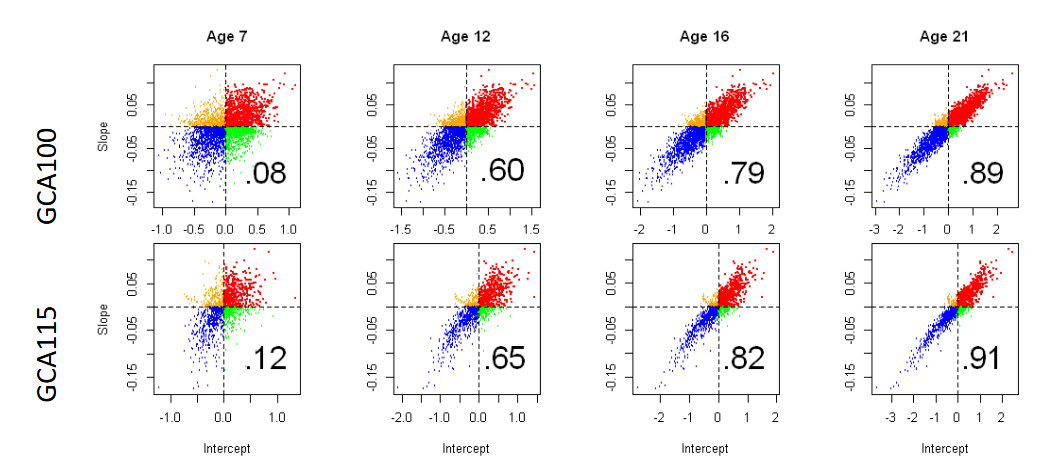
There was a significant sharp increase in the intercept and slope correlation from age 7 to age 21 (Figure 3). At age 7, correlations for both groups were quite low and non-significant (.08 and .12). These outcomes suggest that individuals with the higher mean GCA scores at age 7 were not necessarily those experiencing subsequent meaningful GCA increments. In contrast, individuals with the higher scores at ages 12, 16, and 21, were also those with the larger GCA increments (red points), whereas the individuals with the lower scores at these same ages endured the larger GCA decrements (blue points). The huge correlations at age 21 (.89 and .91) demonstrate the highly heterogeneous experiences for individuals who increased their GCA scores compared with those who decreased their GCA scores.

In summary, the outcomes from the unconditional model suggest a) a meaningful GCA decrement over the studied period, b) remarkable intraindividual variability in the rate of change, and c) a very strong relationship of GCA at age 21 with the rate of change. These findings support underlying potential explanatory factors predictive of the observed change. The conditional model addresses what might be these factors accounting for the interindividual differences on the intraindividual change in GCA (Figure 1).
Conditional GCA model with predictors
The conditional model outcomes shown in Table 3 summarize the associations of the main predictors driving the differences outlined in the unconditional GCA model.
There were two time-varying predictors, home environment (HE) and behavior problems (BP), and six time-invariant predictors, two polygenic scores, SES (age 7), home chaos (age 12), school environment (age 16), and life events (age 21). The evaluation of model fit is shown in the second half of Table S3. The outcomes suggest a slight deterioration in model fit regarding the highly significant chi-square values, and rather low CFI and TLI values. Nonetheless, stringent fit indices such as the RMSEA and SRMR yielded fairly optimal values, suggesting that this model represented the observed data well.
Time varying and time-invariant predictors on GCA intercepts
The top half in Table 3 shows the association of the predictors of change with the intercepts in GCA at ages 7, 12, 16, and 21. For the GCA115 group, time variant predictors were essentially unrelated with GCA intercepts, except for the negative association (-.21) between BP-Intercept and GCA at age 7. This finding was in sharp contrast with the GCA100 group negative associations between the BP-Intercept and BP-Slope with GCA. The relationship between BP-Intercept and GCA decreased from age 7 (-.36) to age 21 (-.17), while being also negative, and remaining more stable, for the BP-slope. Higher BP scores in specific time points (intercept) and in its rate of change (slope), associated with higher decrements in GCA. Moreover, the intercept and slope of HE also associated with GCA at age 12 (.12, .18) and age 16 (.10, .13), indicating that higher HE scores (intercept and slope) related with higher scores in GCA. These outcomes indicate that GCA115 individuals were unaffected by the HE and BP time varying covariates compared with those individuals within GCA100 at these respective ages.
GCA100 and GCA115 were more alike, however, regarding time-invariant predictors. Polygenic scores and SES showed the more robust associations with GCA at each respective age, even though polygenic scores were unrelated with GCA at age 7 for both groups. Higher polygenic scores and higher SES related with higher GCA quite consistently. Home chaos was negatively related with GCA, a relationship that became weaker with increased age. At age 12 and age 16, more disorder at home associated with lower GCA scores. Conversely, a higher school engagement related to higher GCA scores at age 16 and 21. Life events diverged between GCA100 and GCA115. Again, GCA115 were unaffected by life events, whereas for GCA100 more life events related with lower GCA scores at age 16 and age 21. Across age, the coefficients of determination (R2) varied between .15 and .32 for GCA100, and between .20 and .28 for GCA115.
Time varying and time-invariant predictors on GCA slope
The bottom half of Table 3 shows the association of time invariant and time varying predictors with the slope in GCA. The outcomes were virtually the same at the four intercepts (age 7, 12, 16, and 21) because the rate of change within the observed period should be the same regardless of any specific intercept. Regarding time varying predictors and for GCA115, higher BP-Slope associated with more decrements in GCA (-.24 at age 21). This pattern was more robust, however, for GCA100 (-.38 at age 21).
Time-invariant predictors showed more associations with GCA, although with some differences when comparing both groups. Thus, for instance, the Lee 2018 polygenic and SES indices associated more robustly with the rate of change in GCA for GCA115 than for GCA100, whereas the polygenic Okbay 2022 score was more strongly related with the rate of change in GCA for GCA100 than for GCA115. These relationships underline that higher polygenic and SES indices relate with greater positive rates of change in GCA. Similarly, higher scores in school engagement also relate with greater increments in GCA over time, which were higher for GCA115 than for GCA100. Life events exerted a negative impact on GCA for GCA100, whereas these events were unrelated with GCA for GCA115. For GCA100 and GCA115 individuals at age 21, these predictors accounted for 31% and 27% of the variability in GCA, respectively.
Taken together, the conditional model outcomes suggest three conclusions. First, BP had a negative impact on the dynamic change in GCA, although GCA115 individuals were largely unaffected. Second, polygenic scores, SES, and school engagement explained interindividual differences in the intraindividual change in GCA. Finally, the change in GCA was unrelated with home-related variables.
Discussion
Here we have modelled cognitive ability developmental changes applying latent curve models (LCMs). Cognitive ability measures were obtained overtime beginning at age 4 and ending at age 21. Several personal and situational variables were considered to test the hypothesis that if cognitive changes run in tandem with the brain maturational processes relying on the general guidelines provided by the genome, then personal factors (including children’s polygenic scores) would predict the identified cognitive ability changes, whereas situational factors related with their homes and their schools will lose predictive power as the children age.
LCMs treat measures considered across time points (4, 7, 12, 16, and 21 yrs.) as dynamic observed indicators underlying two latent factors, intercept and slope (Figure 1). The intercept factor represents average scores at the start (time 1) or at the final (time n) measurement of the observed time interval, whereas the slope factor represents the rate of change throughout the considered range, from time 1 to time n. After testing the LCMs in two groups of children classified according to their general cognitive ability (GCA) scores at age 7, namely, those scoring on the average range (G100, n = 3,958) and those scoring one standard deviation above the mean (G115, n = 1,580), we obtained the results described earlier and discussed next.
General outlook
The outcomes obtained from the unconditional LCMs suggest a meaningful GCA decrement over the period studied. However, remarkable intraindividual variability in the rate of change was identified. Also, a very strong relationship between GCA at age 21 and the rate of change over time was observed (Figure 3). Taken together, these findings suggest underlying factors that might be predictive of the observed change in GCA. In this later regard, the conditional LCMs addressed what might be these factors accounting for the interindividual differences observed in the intraindividual change. These conditional models provided outcomes suggesting that high cognitive ability individuals were unaffected by the home environment and by their behavior problems assessed across time. Moreover, high ability children were insensitive to their life events, whereas those in the middle range of the cognitive ability distribution experienced a negative impact from these events. Higher scores in school engagement were related with greater increments in GCA across time. Moreover, higher polygenic and SES indices were related with better GCA scores and with greater positive rates of change in GCA (Table 3).
These findings support the conclusion that the followed children and adolescents were cognitively mobile to a substantial degree. Figure 2 reveals that regardless of their general cognitive ability scores at age 7 (between 100 and 115, or greater than 115) most children change their cognitive marks within their respective groups. More specifically, of those belonging to GCA115 at age 7 only 16% kept their high general cognitive ability scores at age 16. Of those belonging to this group at age 12, only 23% kept their high scores. And of those showing high ability at age 16, only 24% kept their high scores. Regarding the group of children showing scores between 100 and 115 at age 7, an 8% moved upwards and obtained high ability scores at age 16, a 6% moved upwards from age 12 to age 16, and an 8% moved also upwards from age 16 to age 21. Therefore, there were remarkable changes in GCA across age.
To use an analogy that may be illustrative, high-level athletes are often appreciated on the understanding that the difficult matter is not to reach a high level but to maintain it for a long time. It seems that this would not apply to high cognitive ability where it is three times more likely to remain at high cognitive ability than to reach it from average levels.
In sum, this observed trend allows us to conclude that: 1) the percentage of subjects in the GCA100 group who raise their GCA level is relatively small and remains stable in all age groups considered, and 2) the percentage of subjects in the GCA115 group who maintain their GCA level is on average 3 times higher, and tends to stabilize from age 12 at around 24 %. Stability values computed after the observed GCA scores were consistent with these movements across the cognitive ability distribution. Thus, for instance, the correlation between GCA scores at age 4 and at age 7 was 0.32, between age 7 and age 12 it was 0.45, between age 12 and age 16 it was 0.59, and between age 16 and age it was 0.65 (Table S1).
Therefore, we can conclude that: 1) it does not seem very useful to detect high cognitive ability at an early age; 2) the age at which it would make sense to identify high cognitive ability would be age 12 (on average), and 3) it would always be necessary to carry out follow-ups to assess the stability of the level achieved.
The Brain Connection
Changes in GCA during this period of the life span were expected. Cognitive ability scores increase across development and reach their peak, on average, at age 16 (Haier et al., 2023; Steinberg, 2015). This process of psychological change runs in tandem with brain maturation. Thus, for instance, studying a representative sample of children and adolescents (from age 6 to age 21) from the Pediatric MRI Data Repository, Roman et al. (2018) analyzed the relationships between intellectual and cortical development. This study modeled the associations between developmental cortical changes and variations in general cognitive ability (g) across time and at the latent level, revealing an average increase in g equivalent to one standard deviation. Therefore, participants turned smarter across the observed period. These cognitive ability changes were correlated with developmental cortical changes, with a greater decrease in cortical values along with smaller increases in cognitive ability over time. These brain changes revealed a highly dynamic nature. Regarding cortical thickness, no changes were observed until age 10, significant thinning was observed for individuals with lower cognitive ability scores in early adolescence, and generalized thinning was identified for all individuals. With respect to cortical surface area, expansion was observed until age 9, adolescents with better cognitive ability scores showed surface contraction at age 12, whereas this contraction process endured until age 16 in those adolescents with lower cognitive scores. The findings suggested that cortical changes should not be generalized to individuals with distinguishable cognitive ability levels. Individuals with better cognitive scores showed cortical thickness preservation, whereas those with lower cognitive scores showed remarkable thinning in early adolescence. These results were also consistent with the developmental research by Burgaleta et al. (2014).
The dynamic nature of these brain-intelligence developmental changes was further corroborated by longitudinal research applying latent change score models to studying children and adolescents (Estrada et al., 2019). Their research question was: what are the time-related associations between cognitive ability changes and cortical structure maturation? The findings revealed that brain and cognitive ability changes related to each other reciprocally. The rate of change, but not previous levels in the brain and cognitive ability indices, was predictive of later changes in both instances.
These examples show that biological maturation relates to brain and cognitive ability changes (Zhou et al., 2015). This relationship is, however, complex and intricate. For instance, a recent theory of educational priorities suggests a developmental three-tier hierarchy of abilities comprising executive and awareness processes, information management, and reasoning, self-evaluation, and knowledge, which consolidate at each of three key developmental stages, preschool, primary education, and secondary education (Demetriou et al., 2023). In this view, the decreasing trajectories of high ability children at age 7 identified here, might suggest that cognitive development may not consolidate well certain abilities at this specific period, which are important for the transition to the next developmental period. Consequently, their ability might have dropped because of difficulties in stabilizing the next time-relevant ability.
Children and adolescents systematically engaged with cognitively demanding activities, for whatever reason, might speed up their spontaneous maturation process. This fastest rate of change might, in turn, change their brains to a significant degree. But if this is the case, why do planned attempts to increase cognitive ability fail to show long lasting effects? The meta-analysis by Protzko (2015) quantified this fadeout effect, suggesting that it can be attributed to the fact that children return to impoverished environments after the cognitive intervention ends. Nevertheless, there might be further explanations and one of them may have to do with how genetic differences drive brain and cognitive ability development.
The latter possibility can be seen as consistent with the demonstrated increased heritability of cognitive ability differences with age (Haworth et al., 2010). Bouchard (2013) referred to this fact as the Wilson effect. He wrote: “contrary to the widespread belief that the slings and arrows of outrageous fortune accumulate over time, the brain appears to be a robust and resilient mechanism (…) the development of cognitive ability is strongly influenced by an intrinsic genetic program that expresses itself differently at different ages (…) cognitive ability appears to be elastic rather than plastic.” The findings reported by Kawakami et al. (2024) regarding educational achievement, and described at the introduction section, seem consistent with this view. Here we found increased predictive validity with age of the analyzed polygenic scores, from 0.08 at age 4 to 0.35 at age 21. This same pattern was identified for SES, with predictive values ranging from 0.14 at age 4 to 0.37 at age 21 (Table S1). Nevertheless, this SES result cannot be clearly interpreted because the index was computed, as noted above, after the educational and occupational credentials of both parents and we know that these variables are genetically charged. When polygenic scores for parents and children are considered, evidence supports the conclusion that the genuine impact of SES, when controlling for the shared genetic factors, is negligible. The research by McGue et al. (2020) concluded that “the offspring-parent difference in measured genetic endowment was significantly correlated with offspring-parent difference in general cognitive ability.” Furthermore, offspring social mobility, based on their own educational and occupational credentials, was unrelated with parents’ social background.
The Question of Why
Scarr (1997) made a systematic comparison of behavior-genetic and socialization theories of intelligence. She concluded that objective environments are generally much less psychologically relevant than knowing how people construct their experiences after these environments. The most important thing to understand from a scientific perspective is “how individuals integrate what they encounter with what they are.” In this same regard, Jensen (1997) focused on the nongenetic variance presumably relevant for general cognitive ability differences, concluding that brains must not be easily shaped by the environment for avoiding compromising their adaptive capacity. This view is consistent with the acknowledged fact that human brains are highly sensitive to their environments. As extensively discussed by Eagleman (2020) human brains dynamically reconfigure themselves for figuring out the changing outer world: “our DNA sets up a dynamic system that continually rewrites its circuitry to reflect the world around it.” This is true, but, as he also acknowledges, brains are not born as blank slates: “fibers from your eye don’t need to learn how to find their targets deep in the brain; they simply follow molecular cues and hit their goal –every time. For all this sort of hardwiring, we can thank our genes.” Can we thank our genes too regarding general cognitive ability? Is the development of cognitive ability mainly guided by what is written in the genome?
Given the demonstrated high heritability of general cognitive ability when individuals reach their adulthood, it is tempting to answer yes to the previous questions. Evidence useful for testing the causal assumptions embraced by current psychometric models of cognitive ability supports the assumption that specific cognitive abilities can be highly sensitive to the impact of environmental (nongenetic) factors, whereas general cognitive ability (g) is not. Protzko and Colom (2021) reviewed these psychometric models considering what we already know regarding the positive effects of cognitive training, along with the negative effects of focal brain lesions, over cognitive abilities. They concluded that these effects are local instead of global, meaning that they do have an influence over specific cognitive abilities but are not reflected on the higher-order general common factor. The psychometric model that can accommodate better these findings shows a bifactorial structure, meaning that measured cognitive performance differences are causally influenced by specific cognitive abilities, such as verbal or visuospatial abilities, and by the higher-order factor representing general cognitive ability (g). As shown by the genetically sensitive study of Panizzon et al. (2014) the general factor of intelligence is highly heritable (86 %). Their findings supported the view that general cognitive ability is a genetic phenomenon (Plomin & Spinath, 2002).
Implications for Practice
Given the substantial developmental changes in general cognitive ability scores reported in the present study (Figure 2), there is an important question demanding an answer: should we trust early identification of high cognitive ability children or should be encourage systematic evaluation across childhood and adolescence?
We have seen that the number of children showing high cognitive ability in early childhood and preserving their scores as they age is low. Therefore, it is now a good moment to remember the cautionary note raised by Asbury and Plomin (2014). As seen at the introduction section, static classifications are largely unwarranted. Those showing high cognitive ability at early childhood might lose their marks as they age. Conversely, those showing average cognitive ability may increase their marks as they age, although this is much less likely, as seen above. Moreover, the large variability, and remarkable decrease in the trajectories of children identified as high in cognitive ability at age 7, bears an important connection with educational policy. Nonetheless, the findings reported here underline a stronger impact of genes than of school engagement on the change in cognitive ability. As suggested elsewhere, therefore, the importance of genes for learning implies that some children are better able to learn than others, supporting the need of models centered on the individual rather than on the general population (Asbury & Plomin, 2014; Demetriou et al., 2023).
The straightforward implication of the evidence found in the present study is, therefore, that systematic evaluation is highly recommended, especially at the critical age of 12 years. The educational system must avoid trusting in premature evaluations of pupils’ general cognitive ability and the attached administrative labels. Remarkable changes in the relative standing of children and adolescents regarding their general cognitive ability should be expected. Therefore, in contrast with the diminution of rigorous assessment of cognitive abilities in modern educational systems (Murray, 2008) and in the light of the current findings, planned systematic follow ups are strongly recommended to avoid unjustified educational actions.
Limitations
The TEDS dataset is certainly comprehensive, including a number of personal and situational factors measured since early childhood and using a prospective design. These factors comprise intelligence, personality, self-efficacy, health, wellbeing, home environment, school environment, academic achievement, behavior problems, and polygenic scores of a set of relevant psychological variables. Nevertheless, TEDS did not register data regarding the brains of their participants, even though neuromarkers might be of great help for enhancing our understanding of the notable developmental changes reported in the present study. There are several candidates as neuromarkers distinguishing high and normative cognitive ability individuals (Santonja et al., 2021a, 2021b) that might contribute to identify those individuals losing their high cognitive marks across their development. Finding the connection in the brain between the genomic and the environmental relevant signals requires obtaining the structural and functional features of potential interest. The ENIGMA Consortium is an example of this recommended approach (Thompson et al., 2020).
Still another limitation of the present study is that we were unable to analyze all the potentially relevant processes of change because of the technical requirements of the latent curve models considered here (Preacher et al., 2008). At least three equivalent measures are needed, but these were unavailable in some instances.
Concluding Remarks
Here we have documented what it is, not what it could be (Plomin, 2018) after the analyses of prospective longitudinal data carefully collected through the TEDS initiative. As metaphorically stated by Belsky et al. (2020) doing this research is “like being in one of those nature documentaries where the lion stalks the gazelle; the scientists never try to save the gazelle or stop the lion, they just observe.” We have shown that children with high general cognitive ability are largely mobile as they age. Of those scoring one standard deviation above the mean at age 7, only a tiny minority preserve their high ability marks afterwards. Moreover, some children scoring below the standards of high ability at age 7 move upwards in the cognitive distribution, although this is three times less likely than maintaining the standard of high cognitive ability for those who already exhibited it.
This led us to conclude that early identification of high ability children should not be trusted and, therefore, planned follows up must be mandatory to avoid unwarranted requirements in educational settings and beyond. The movements across the cognitive distribution were mainly predicted by children’s polygenic scores, along with the educational and occupational credentials of their parents, especially since age 12. The measures of home environment, behavior problems, and life events were generally unrelated with these cognitive ability changes. And this is especially true for individuals with high cognitive ability. Therefore, personal factors (nature) seem more relevant than situational factors (nurture) to predict cognitive ability changes across childhood and adolescence.
Acknowledgements
We gratefully acknowledge the ongoing contribution of the Twins Early Development Study (TEDS) participants and their families.
TEDS is funded by a UK Medical Research Council (MRC) program grant (MR/V012878/1) to TC Eley (previously MR/M021475/1 to R Plomin).
This research was performed within the Catalonian Consolidated Research Group 2021SGR 01423.